 -
Volume 47,
Issue 2,
1967
-
Volume 47,
Issue 2,
1967

Volume 47, Issue 2, 1967
- Articles
-
-
-
Purine-Requiring Auxotrophs of Coprinus lagopus (sensu Buller)
More LessThe allelic relationships between a series of independently-induced adenine mutants of Coprinus lagopus were investigated, and limited complementation maps of three of the loci constructed. By means of nutritional tests and a search for aminoimidazole accumulants in the mycelium the adenine loci were correlated with steps in the purine biosynthetic pathway. Four loci can be assigned to (unknown) steps preceding closure of the imidazole ring, two others to specific steps occurring after closure of the ring. One of these latter represents a deficiency in the enzyme adenylosuccinase.
-
-
-
-
Lipolytic Activity by Oral Pleuropneumonia-Like (Mycoplasma) Organisms
More LessNinety-five strains of Mycoplasma were isolated from human saliva; all were micro-aerophilic; 72 were actively lipolytic and 23 were non-lipolytic and were inhibited by Tweens. A type culture of human type 4 (Mycoplasma salivarum) was culturally identical with the lipolytic strains, and two type cultures of M. orale with the non-lipolytic strains. The strains serologically identifiable as M. orale were non-lipolytic, whereas those identifiable as M. salivarium were of both types. Comparison was also made with type strains from other sources. Those of human origin were non-lipolytic and those of saprophytic or animal origin were slightly lipolytic.
-
-
-
The Utilization of Propionate by Micrococcus denitrificans
More LessSuspensions of Micrococcus denitrificans, growing with propionate as sole carbon source, incorporated 14C from [1-14C]propionate or from sodium [14C]bicarbonate, initially into succinate and then into intermediates of the tricarboxylic acid cycle and amino acids derived therefrom. In the presence of 4 mM-sodium arsenite, the oxidation of propionate and of L-malate by washed organisms proceeded only to the level of pyruvate, which accumulated: when sodium [14C]bicarbonate was also present, the pyruvate formed from propionate, but not that formed from L-malate, was highly radioactive. Cell-free extracts of propionate-grown M. denitrificans catalysed the formation of labelled methylmalonyl-coenzyme A, succinyl-coenzyme A and succinate from sodium [14C]carbonate+ ATP + either propionate and coenzyme A, or propionyl-coenzyme A. The evidence thus obtained indicates that propionate enters the tricarboxylic acid cycle of M. denitrificans, preponderantly via activation to propionyl-coenzyme A, followed by carboxylation to methyl-malonyl coenzyme A, isomerization to succinyl-coenzyme A and hydrolysis to succinate.
-
-
-
A Kinetic Study of the Mode of Growth of Surface Colonies of Bacteria and Fungi
More LessA model for the growth of microbial colonies on the surface of a solid nutrient medium is discussed. The model accounts for the constant rate of increase in the colony radius which is characteristic of a fungal colony growing on the surface of a nutrient medium.
Experiments showed that bacterial colonies after about 12 hr of development showed a virtually constant rate of radial growth over a 12 hr period. Over longer periods (24 hr) a gradual decline in the colony radial growth rate was apparent. The initial rate of radial growth of the bacterial colony was a useful parameter of the growth rate of the organism. The effects on the initial colony radial growth rate of the following factors were determined: initial nutrient concentration depth of agar layer; maximum specific growth rate (In 2/minimum doubling time); oxygen partial pressure; humidity of gas phase; temperature. Three bacterial types, Escherichia coli, Kleb-siella aerogenes and Streptococcus faecalis were studied. With E. coli growing in minimal medium in air at 1 atm. pressure when the growth was glucose-limited, oxygen became a limiting factor when the glucose concentration exceeded 0.25% (w/v). With a glucose concentration of 1% (w/v), the growth was strongly inhibited, probably by toxic products
When the colony growth was glucose-limited and oxygen was present in excess, the relation between initial colony radial growth rate (K r), the initial glucose concentration (s o) and the maximum specific growth rate (αm) was K r = k 2(√s 0 − √S i) √αm
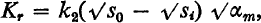
where k 2 is a constant; s i, called the ‘lag concentration’, is a value of the glucose concentration which must be exceeded before growth of the colony can occur. The value of s i was very small or negligible except with a certain type of inhibitory condition, such as an over-optimal concentration of oxygen, which could be overcome by the organism's metabolic activity. Direct proportionality between K r and √αm was found by varying the maximum specific growth rate by adding sulphanilamide. When αm was varied by temperature changes the linear relation between K r and √αm did not hold. The implications of these results and their potential applications are discussed.
-
-
-
Enzyme and Permeability Changes During Morphogenesis of Nocardia corallina
More LessO. R. Brown and S. RedaInitial growth of Nocardia corallina occurred by germination and coenocytic hyphal development which resulted in a large surface-to-volume increase. During this period, endogenous respiration and the ability to oxidize fructose increased rapidly, but the stimulation of oxygen uptake by glucose decreased to zero. This inability to utilize glucose was due to loss of permeability rather than lack of a specific enzyme required for glucose metabolism. This was established by the following data: (1) cell-free extracts of hyphae contained the enzymes for conversion of glucose into fructose-6-phosphate; (2) intact hyphae oxidized fructose; (3) incubation of hyphae with cetyltri-methylammonium bromide increased cell permeability to glucose and oxygen uptake was stimulated. Comparative determinations of enzymes involved in the early steps of glucose metabolism showed quantitative changes associated with the period of impermeability to glucose. The loss of permeability to glucose may represent an initial metabolic change fundamental to morphogenesis in this microbe.
-
-
-
On the Taxonomic Status of ‘Quin’s Oval’ Organisms
More LessPurified suspensions of the sheep rumen organism known colloquially as ‘Quin's oval’ were prepared from sheep rumen liquor. The chemical composition of cell walls prepared from the organisms of these suspensions was found to include protein, lipid and polysaccharide. A fraction from these walls, obtained after removal of much of the protein and polysaccharide, was shown to contain muramic acid, glucosamine and diaminopimelic acid. These findings are consistent with this organism being regarded as a Gram-negative bacterium.
-
-
-
The Morphology and Ultrastructure of the Spore and Exosporium of Some Clostridium Species
More LessThe spores of original isolates of Clostridium species obtained from fresh and processed marine fish and from marine bottom deposits were compared with the spores of culture collection strains of the same species. These strains were Clostridium botulinum type E, C. sporogenes, C. bifermentans and C. sordellii. Spores of unclassified, non-toxigenic isolates which resemble C. botulinum type E, of ‘OS mutants’, of culture collection strains of C. botulinum types A and B and an isolate of type F were also examined. The gross morphology and ultrastructure of the spores and exosporia were studied in electron microscopic preparations by using the techniques of metal shadowing, carbon replica, negative-staining and ultra-thin sectioning. Spores of some strains were found to possess tubular appendages. The morphology of the spores and the ultrastructural detail of the spore appendages and exosporia are described.
-
-
-
Studies on the Virulence of Hospital Strains of Pseudomonas aeruginosa
More LessThe virulence of 36 strains of Pseudomonas aeruginosa was studied by intravenous injection into mice; the strains were found to fall into three broad groups, of high, medium and low virulence. This difference could not be related to any biochemical property studied, pyocine type nor antibiotic sensitivity pattern. It was found that 24 of the 30 strains of the high and medium virulence group produced large colonies on agar while 5 of the 6 strains of the low virulence group produced small colonies when plated under similar conditions. To test this observation a further series was collected and the strains were allocated to the high or low virulence category on colonial appearance. Mouse challenge with these fresh strains showed that the prediction was accurate. The large colony type had a faster growth rate on agar and in broth than the small. When a strain was grown in agar it was found to be more virulent than when harvested from broth (P = 0.02–0.01). Chromatography studies on slime derived from all three categories showed that they were apparently chemically similar. The amount of slime produced by a standard number of bacteria was measured for agar and broth grown cultures. The yields for the two media were approximately the same with large-colony forms. Small-colony forms produced similar amounts to the large forms on plates, but up to 15 times as much when grown in broth.
The ability to kill mice was derived in part from early toxic death; the remainder died of renal disease. Study of the initiation of kidney infection showed that large-colony types were more successful in maintaining their numbers in the kidneys over the first 5 hr. Strains grown on nutrient agar did not give higher counts in the kidney than those from broth in the first 5 hr but after 24 hr they grew faster, infected more kidneys and killed more mice.
-
-
-
Chemical Composition of Hyphal Wall of Phycomycetes
More LessIsolated hyphal walls from Phytophthora heveae, Pythium butleri and Saprolegnia ferax were hydrolysed and compared in quantitative composition. A high carbohydrate content was found in all these walls (80–90%); also a small amount of glucosamine (1–2%), lipids (1–2.5%) and protein. Quantitatively the cell walls differed appreciably in protein (1–11%). Five neutral sugars (glucose, galactose, mannose, rhamnose, ribose) were detected by chromatography; glucose was always the main monosaccharide. From the results of acid-extraction of walls and enzymic hydrolysis it was concluded that a portion of cell-wall polysaccharide was cellulose (30–45%). The amino acids found were those typical of normal protein hydrolysates and containing also hydroxyproline.
-
-
-
Growth and Characterization of Nocardiophages for Nocardia canicruria and Nocardia erythropolis Mating Types
More LessTwo nocardiophages were isolated from soil samples: one, called ⊘C, was specific for the Nocardia canicruria mating type; the other, called ⊘C, proliferated in N. canicruria and N. erythropolis mating types. Inactivation of phage ⊘C was rapid in NaCl solutions, with chloroform, 0–3% hydrogen peroxide and 30% (v/v) ethanol in water. Phage ⊘C behaved similarly but was less labile in NaCl solutions. Both phages were resistant to diethyl ether. Phage ⊘C could be propagated to titres of 1010 plaque-forming units/ml. on a medium containing peptone, yeast extract and calcium nitrate; the highest observed titres of phage ⊘C were also produced in this medium. A chemically defined medium for proliferation of phage ⊘C consisted of inorganic salts, valine, isoleucine, leucine, nicotinamide, glycerol. Other variables which affected the growth of these phages were the amount of phage and host inocula and the age of the host before infection. Phage ⊘C was distinct from other nocardiophages in its ability to attach quickly and efficiently to its host; 95% of the phage attached, and attachment was time and temperature dependent. Attachment of phage ⊘EC was slow and inefficient, reaching only 34%. The latent period for phage ⊘C was about 25 min., the increase period was 35 min. and the burst size about 60 particles/infective centre. The latent period of phage ⊘EC was 180 min. and the burst size about 20 particles. Both these nocardiophages possessed the typical actinophage morphology.
-
-
-
Chemical and Electron Microscope Studies on Fractions Prepared From Coats of Bacillus Spores
More Less‘Coat fractions’ were prepared from spores of six Bacillus species by mechanical disruption, centrifugation, and digestion with lysozyme. The coats were successively extracted with dilute NaOH, treated sonically (mildly), and digested with the proteolytic enzyme preparation Pronase. Bacillus megaterium QM B1551 was studied in detail. Electron microscopic studies were made at the various stages. Three fractions were obtained and partly characterized chemically. The alkali-soluble fraction consisted mainly of protein(s). The paracrystal fraction solubilized by sonic treatment consisted mainly of protein with a chemical composition and physical characteristics similar to those of keratins. The final resistant residue fraction of the coats after hydrolysis contained amino acids, phosphorus and muramic acid, and may be composed of a phosphomuramyl polymer to which a peptide and/or protein is linked. Electron microscopic observation suggested that the spore coat of B. megaterium QM B1551 consisted of at least three components: a middle, paracrystal fraction is sandwiched between or ‘cemented’ with the alkali-soluble fraction on one side and the resistant residue layer on the other. Large differences were found in the composition of the coats and in the P contents of the resistant residue of the various organisms.
-
-
-
Differentiation of Strains of Staphylococcus epidermidis Isolated From Bovine Udders
More LessStrains of Staphylococcus epidermidis isolated from bovine udders were differentiated by comparing biochemical tests, serological typing of proteolytic enzymes, colonial morphology, and the spectrophotometric analysis of pigments extracted with methanol. The cultures were divided into proteinase-positive and proteinase-negative groups, based on their proteolytic activity on skim milk agar, staphylococcus medium no. 110 (Difco), and gelatin. Most of the proteinase-negative cultures produced acetoin, whereas the opposite was true for the proteinase-positive cultures. A further subdivision of the cultures in each group was made by using Baird-Parker's (1963) biochemical subgrouping scheme. The proteinase-positive cultures were also subdivided by serological typing of their proteolytic enzymes into five groups, B, F, G, H, and NR, a non-reacting group. Approximately three-quarters of the cultures in the first four proteinase groups could be placed in one of Baird-Parker's biochemical subgroups. The cultures in the NR group, however, were sufficiently distinctive in their biochemical reactions to be considered as a separate biochemical subgroup. The colonies formed by the cultures were classified into five types, each consisting of two to four forms. No absolute relationship was found between the spectral absorption curves of the pigments and proteinase groups, although there were differences in the types of absorption curves that predominated in each proteinase group. Nine cultures which produced coagulase were more closely related to S. epidermidis than to S. aureus in that they did not utilize mannitol anaerobically or produce α-or β-type haemolysis, they were non-pigmented and were less active biochemically. Also, serological typing of the proteolytic enzymes of three of these cultures resulted in one being classified as group G and two as group H. Group H contained only these two cultures. In contrast 27 cultures of S. aureus were classified as group A. Except for coagulase production, the biochemical reactions and the serological grouping of the proteolytic enzymes suggested a classification of S. epidermidis for the nine cultures.
-
-
-
A Solid Medium Test for Measuring Growth Inhibition and Neutralization of Mycoplasma mycoides by Immune Bovine Serum
More LessThe growth of Mycoplasma mycoides var. mycoides was inhibited and neutralized by immune serum from cattle which had recovered from infection with M. mycoides. Although mycoplasmal neutralization is not necessarily the same as virus neutralization, it is suggested that this terminology be retained since it is accepted and is descriptively appropriate. The effect of immune serum on colony growth was determined by incubating test serum in plastic cylinders on nutrient agar with pre-incubated microscopic M. mycoides colonies. Optimal inhibition and neutralization occurred when serum and colonies were incubated at 30° before final incubation at 37°. Inhibition of growth and neutralization also occurred at 37° and 21° inhibition but no neutralization occurred at 5°.
-
-
-
Isolation, Classification and Nutritional Requirements of Cellulolytic Cocci in the Sheep Rumen
More LessTwenty-five strains of cellulolytic cocci were isolated from the rumen contents of seven sheep, over a period of two years. Six sheep were maintained at roughly constant body weight on chaffed lucerne hay or a lucerne chaff/oat chaff mixture. One strain was obtained from an animal on a low protein diet.
The bacteria were classified as Ruminococcus albus or R. flavifaciens by the accepted monothetic classification. Morphological, cultural, nutritional and fermentation data were then used to classify them by a numerical method. It was concluded that the numerical classification of these strains agreed closely with the classification by more established methods, but justified the assignment of relationship between members of the same species.
The nutritional characteristics of 11 strains capable of growth in partially defined medium were investigated. An organic nitrogen source was essential for three strains; the remaining eight strains grew better with casein hydrolysate than with ammonium sulphate as the sole nitrogen source. Although ammonium ions were not essential for growth, only three strains were capable of growth in the absence of volatile fatty acids. CO2 was essential for 10 of 11 strains.
-
-
-
Antigenic Relations of Cellulolytic Cocci in the Sheep Rumen
More LessTwenty-five strains of ruminococci were isolated from seven sheep fed chaffed lucerne hay, or mixtures of cereal chaff and lucerne chaff, for 18 months. Antiserum to each strain was produced in rabbits and used to classify the strains by cross-agglutination. Two main groups and a subgroup were identified which served to characterize half (12 of 25) of the strains isolated. Group 1 (7 strains) and the subgroup (2 strains) were all classified as Ruminococcus albus. Group 2 comprised 3 strains of R. flavifaciens. An indirect fluorescent-antibody technique was also used to show antigenic relationships amongst the isolates. Fluorescence cross-reactions were obtained among 13 strains of R. albus using group 1 antisera. Similarly a group 2 antiserum labelled 7 of the 12 strains of R. flavifaciens isolated. The persistence of these serotypes in the rumen was demonstrated by the isolation of cross-agglutinating strains of R. albus from one animal at intervals throughout the experimental period. Recovery of cross-agglutinating strains of R. albus and R. flavifaciens from more than one animal suggests that a given serotype becomes disseminated among animals which are housed together. Furthermore, the isolation of group 1 and group 2 ruminococci from the same sheep indicates that the group-specific determinants were not dependent on the rumen environment for expression. It is concluded that selection was rigorous enough to maintain recognizably different genetic strains in the rumen against mutation pressure.
-
- Books Received
-
- Corrigenda
-
Volumes and issues
-
Volume 170 (2024)
-
Volume 169 (2023)
-
Volume 168 (2022)
-
Volume 167 (2021)
-
Volume 166 (2020)
-
Volume 165 (2019)
-
Volume 164 (2018)
-
Volume 163 (2017)
-
Volume 162 (2016)
-
Volume 161 (2015)
-
Volume 160 (2014)
-
Volume 159 (2013)
-
Volume 158 (2012)
-
Volume 157 (2011)
-
Volume 156 (2010)
-
Volume 155 (2009)
-
Volume 154 (2008)
-
Volume 153 (2007)
-
Volume 152 (2006)
-
Volume 151 (2005)
-
Volume 150 (2004)
-
Volume 149 (2003)
-
Volume 148 (2002)
-
Volume 147 (2001)
-
Volume 146 (2000)
-
Volume 145 (1999)
-
Volume 144 (1998)
-
Volume 143 (1997)
-
Volume 142 (1996)
-
Volume 141 (1995)
-
Volume 140 (1994)
-
Volume 139 (1993)
-
Volume 138 (1992)
-
Volume 137 (1991)
-
Volume 136 (1990)
-
Volume 135 (1989)
-
Volume 134 (1988)
-
Volume 133 (1987)
-
Volume 132 (1986)
-
Volume 131 (1985)
-
Volume 130 (1984)
-
Volume 129 (1983)
-
Volume 128 (1982)
-
Volume 127 (1981)
-
Volume 126 (1981)
-
Volume 125 (1981)
-
Volume 124 (1981)
-
Volume 123 (1981)
-
Volume 122 (1981)
-
Volume 121 (1980)
-
Volume 120 (1980)
-
Volume 119 (1980)
-
Volume 118 (1980)
-
Volume 117 (1980)
-
Volume 116 (1980)
-
Volume 115 (1979)
-
Volume 114 (1979)
-
Volume 113 (1979)
-
Volume 112 (1979)
-
Volume 111 (1979)
-
Volume 110 (1979)
-
Volume 109 (1978)
-
Volume 108 (1978)
-
Volume 107 (1978)
-
Volume 106 (1978)
-
Volume 105 (1978)
-
Volume 104 (1978)
-
Volume 103 (1977)
-
Volume 102 (1977)
-
Volume 101 (1977)
-
Volume 100 (1977)
-
Volume 99 (1977)
-
Volume 98 (1977)
-
Volume 97 (1976)
-
Volume 96 (1976)
-
Volume 95 (1976)
-
Volume 94 (1976)
-
Volume 93 (1976)
-
Volume 92 (1976)
-
Volume 91 (1975)
-
Volume 90 (1975)
-
Volume 89 (1975)
-
Volume 88 (1975)
-
Volume 87 (1975)
-
Volume 86 (1975)
-
Volume 85 (1974)
-
Volume 84 (1974)
-
Volume 83 (1974)
-
Volume 82 (1974)
-
Volume 81 (1974)
-
Volume 80 (1974)
-
Volume 79 (1973)
-
Volume 78 (1973)
-
Volume 77 (1973)
-
Volume 76 (1973)
-
Volume 75 (1973)
-
Volume 74 (1973)
-
Volume 73 (1972)
-
Volume 72 (1972)
-
Volume 71 (1972)
-
Volume 70 (1972)
-
Volume 69 (1971)
-
Volume 68 (1971)
-
Volume 67 (1971)
-
Volume 66 (1971)
-
Volume 65 (1971)
-
Volume 64 (1970)
-
Volume 63 (1970)
-
Volume 62 (1970)
-
Volume 61 (1970)
-
Volume 60 (1970)
-
Volume 59 (1969)
-
Volume 58 (1969)
-
Volume 57 (1969)
-
Volume 56 (1969)
-
Volume 55 (1969)
-
Volume 54 (1968)
-
Volume 53 (1968)
-
Volume 52 (1968)
-
Volume 51 (1968)
-
Volume 50 (1968)
-
Volume 49 (1967)
-
Volume 48 (1967)
-
Volume 47 (1967)
-
Volume 46 (1967)
-
Volume 45 (1966)
-
Volume 44 (1966)
-
Volume 43 (1966)
-
Volume 42 (1966)
-
Volume 41 (1965)
-
Volume 40 (1965)
-
Volume 39 (1965)
-
Volume 38 (1965)
-
Volume 37 (1964)
-
Volume 36 (1964)
-
Volume 35 (1964)
-
Volume 34 (1964)
-
Volume 33 (1963)
-
Volume 32 (1963)
-
Volume 31 (1963)
-
Volume 30 (1963)
-
Volume 29 (1962)
-
Volume 28 (1962)
-
Volume 27 (1962)
-
Volume 26 (1961)
-
Volume 25 (1961)
-
Volume 24 (1961)
-
Volume 23 (1960)
-
Volume 22 (1960)
-
Volume 21 (1959)
-
Volume 20 (1959)
-
Volume 19 (1958)
-
Volume 18 (1958)
-
Volume 17 (1957)
-
Volume 16 (1957)
-
Volume 15 (1956)
-
Volume 14 (1956)
-
Volume 13 (1955)
-
Volume 12 (1955)
-
Volume 11 (1954)
-
Volume 10 (1954)
-
Volume 9 (1953)
-
Volume 8 (1953)
-
Volume 7 (1952)
-
Volume 6 (1952)
-
Volume 5 (1951)
-
Volume 4 (1950)
-
Volume 3 (1949)
-
Volume 2 (1948)
-
Volume 1 (1947)
Most Read This Month
