 -
Volume 137,
Issue 5,
1991
-
Volume 137,
Issue 5,
1991

Volume 137, Issue 5, 1991
- Physiology And Growth
-
-
-
Catabolite repression by galactose in overexpressed GAL4 strains of Saccharomyces cerevisiae
More LessT. Lodi, C. Donnini and I. FerreroCatabolite repression by galactose was investigated in several strains of Saccharomyces cerevisiae grown on different carbon sources. Galactose repressed as much as glucose; raffinose was less effective. Full derepression was achieved with lactate. The functions tested were l-lactate ferricytochrome c oxidoreductase, NAD-glutamate dehydrogenase, and respiration. Galactose repression was observed only in the GAL4 but not in the gal4 strain. The presence of multiple copies of the GAL4 gene enhanced the repression by galactose. Different alleles of the GAL4 gene and the copy number did not affect glucose repression.
-
-
-
-
Antigenic cell wall mannoproteins in Candida albicans isolates and in other Candida species
More LessPolyclonal antibodies (pAbs) and monoclonal antibodies (mAbs), raised against mannoprotein components from Candida albicans ATCC 26555 (serotype A) blastoconidia and mycelial cell walls, were used to investigate antigenic similarities among wall mannoproteins from other C. albicans serotype A and B strains, and from C. tropicalis and C. guilliermondii. Radioactively labelled walls isolated from cells grown at either 28 °C or 37 °C were digested with a β-glucanase complex (Zymolyase 20T) to release cell-wall-bound mannoproteins. Numerous molecular species with different electrophoretic mobilities were released from the various isolates. Differences appeared to be related to both the organism and the growth temperature. Among the major protein components solubilized were mannoproteins larger than 100 kDa (high molecular mass mannoproteins), heterogeneous in size in most cases. Antigenic homology was detected among the cell wall high molecular mass mannoproteins of the two C. albicans serotype A isolates, whereas significant qualitative and quantitative differences were detected between serotype A and serotype B cell-wall-bound antigenic profiles. Moreover, C. tropicalis and C. guilliermondii wall antigenic determinants were not recognized by the preparations of pAbs and mAbs raised against C. albicans walls. A mannoprotein with a molecular mass of 33-34 kDa was present in the enzymic wall digests of all the organisms studied. When probed with pAbs raised against the protein moiety of the 33 kDa cell wall mannoprotein of Saccharomyces cerevisiae, antigenic cross-reactivity was observed in all cases except C. tropicalis. There appear to be significant antigenic differences between the mannoproteins of different isolates of C. albicans, and between those of C. albicans and other Candida species.
-
-
-
A new pathway for isonicotinate degradation by Mycobacterium sp. INA1
More LessAn organism able to utilize isonicotinate aerobically as the sole source of carbon, nitrogen and energy was isolated from soil samples and identified as a Mycobacterium sp. No growth occurred on any other heteroaromatic substrate tested, except 2-hydroxyisonicotinate, which was shown to be an intermediate in isonicotinate metabolism. Degradation of isonicotinate involved two consecutive hydroxylations leading to 2,6-dihydroxy-isonicotinate (citrazinate). The reactions were catalysed by two different dehydrogenases, which were apparently molybdenum-dependent. Citrazinate was activated by CoA via a synthetase reaction, and the resulting citrazyl-CoA ester was reduced by a NADPH-dependent reductase. In the presence of 1 mm-arsenite, cells accumulated 2-oxoglutarate during isonicotinate metabolism.
-
-
-
The possible involvement of trypsin-like enzymes in germination of spores of Bacillus cereus T and Bacillus subtilis 168
More LessGermination of spores of Bacillus cereus T and Bacillus subtilis 168 was inhibited by the trypsin inhibitors leupeptin and tosyllysine chloromethyl ketone (TLCK) and by the substrates tosylarginine methyl ester (TAME), benzoyl-l-arginine-P-nitroanilide (L-BAPNA) and D-BAPNA. Potencies of these inhibitory compounds were estimated by finding the concentration which inhibited 50 % germination (ID50), as measured by events occurring early (loss of heat resistance), at an intermediate stage [dipicolinic acid (DPA) release], and late in germination (decrease in optical density). In B. cereus T, all the compounds inhibited early and late events with the same ID50. In B. subtilis, TAME inhibited early and late events at the same ID50, but all other inhibitors had a lower ID50 for late events than for early events. This suggests that a trypsin-like enzyme activity is involved at two sequential stages in the germination of B. subtilis spores, one occurring at or before the loss of heat resistance and one at or before the decrease in optical density. Different trypsin-like activities were detected in broken dormant spores and germinated spores of B. cereus T and in germinated spores of B. subtilis by means of three chromogenic substrates: benzoyI-l-phenylalanyl-l-valyl-l-arginine-p-nitroaniIid (L-PheVA), l-BAPNA and d-BAPNA. Separation of extracts of germinated spores on non-denaturing polyacrylamide gels showed that in both species the substrates were hydrolysed by three distinct enzymes with different electrophoretic mobilities. The three enzymes had different Ki values for the above inhibitors. The possibility that l-BAPNase from B. cereus T might be involved in the initial germination steps was suggested by the similarity of ID50 values for germination and Ki values for inhibition of the enzyme in vitro by TAME and TLCK, and by the fact that both germination and the l-BAPNase were reversibly inhibited by TLCK.
-
-
-
Production of type 5 capsular polysaccharide by Staphylococcus aureus grown in a semi-synthetic medium
More LessThe concentration of the type 5 capsular polysaccharide (CP) antigen of Staphylococcus aureus can be measured directly in cultures or cell suspensions by a two-step inhibition enzyme-linked immunosorbent assay (ELISA), using monoclonal antibodies. CP was synthesized during growth on a variety of carbon substrates and its production was not affected by the nature of the carbon source. High levels of yeast extract inhibited CP formation. CP was synthesized in batch culture at the same rate during exponential growth as in the post-exponential phase. Post-exponential CP production contributed at least half the final amount of CP measured. This phenomenon was observed in different culture media, although the specific yield of polysaccharide varied from one medium to another. Post-exponential CP production was observed in the pH range 6–7, but not at pH 8. Post-exponential production was strictly dependent on oxygen availability and did not occur under anaerobic conditions.
-
-
-
Determination of effector molecules in L-arabinose-induced bulge formation and lysis of Escherichia coli IFO 3545
More LessL-Ribulose 5-phosphate (L-Ru5P) was identified as the primary effector molecule of L-arabinose-induced bulge formation in Escherichia coli IFO 3545 observed in nutrient broth with 5 % (w/v) sodium chloride. Hyperinduction of L-arabinose isomerase was due to exogenous sodium chloride and the resulting alteration in the balance of the L-arabinose-metabolizing enzymes resulted in accumulation of L-Ru5P. L-Ru5P induced the lysis of an L-arabinose-negative, L-Ru5P 4-epimerase-less mutant, ara-207, even when directly added to the medium but was not active against the wild-type strain. Some L-arabinose-utilizing (L-arabinose-resistant) revertants of ara-207 were still sensitive to L-Ru5P, indicating the involvement of another mutation in L-Ru5P-sensitivity other than genetic lack of L-Ru5P 4-epimerase. Among the various pentose phosphate esters tested, only L-Ru5P could induce lysis of ara-207. The lytic activity of L-Ru5P was attributed to its effect on bacterial sugar nucleotide metabolism which caused secondary accumulation of uridine 5’-diphosphate galactose (UDPGal), which provoked lysis induction.
-
-
-
Gas vesicle collapse by turgor pressure and its role in buoyancy regulation by Anabaena flos-aquae
More LessFilaments of Anabaena flos-aquae lost over half of their gas vesicles when exposed to light of high irradiance (> 115 μmol m–2 s–1) for long periods (up to 23 h). An investigation using different irradiances showed that the major loss of gas vesicles occurred after prolonged exposure to irradiances exceeding 80 μmol m–2 s–1. In a time-course experiment it was found that 56% of the gas vesicles were lost after 16 h exposure to an irradiance of 135 μmol m–2 s–1. Most of this loss could be accounted for by collapse under turgor pressure, which rose by 0·24 MPa to 0·54 MPa after 16 h. A further unexplained loss of 15 % after 23 h exposure may indicate that prolonged high irradiance can cause weakening of gas vesicles in the cells. In this time-course experiment 98% of the filaments lost their buoyancy after 6 h exposure, before any gas vesicles were lost; this buoyancy change was quantitatively accounted for by the rise in cell carbohydrate. The relative importance of gas vesicle collapse and carbohydrate accumulation as mechanisms of buoyancy loss is discussed in relation to vertical movements of planktonic cyanobacteria in lakes.
-
-
-
Separating the efflux and influx components of net nitrate uptake by Synechococcus R2 under steady-state conditions
More LessN isotopic discrimination was used to determine the ratio of influx to efflux in experiments in which
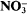 reduction was decreased (and also net
reduction was decreased (and also net 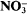 uptake, since under steady-state conditions, net
uptake, since under steady-state conditions, net 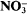 uptake is equal to the rate of reduction) by tungstate. Exponential growth rate constants for the cyanobacterium Synechococcus sp. R2 grown in media containing 0·18 μM-molybdate decreased linearly (0·040 to 0·013 h–1) with increasing tungstate concentration (0 to 0·2 mm). Values of the overall, observed N isotope effect (β
obs, the ratio of the rate of reaction of 14N- to 15N-bearing molecules, normalized to adjust for unequal concentrations) varied inversely and were linear with exponential growth rate. At high tungstate concentration, when the exponential growth rate approached zero (as a result of negligible
uptake is equal to the rate of reduction) by tungstate. Exponential growth rate constants for the cyanobacterium Synechococcus sp. R2 grown in media containing 0·18 μM-molybdate decreased linearly (0·040 to 0·013 h–1) with increasing tungstate concentration (0 to 0·2 mm). Values of the overall, observed N isotope effect (β
obs, the ratio of the rate of reaction of 14N- to 15N-bearing molecules, normalized to adjust for unequal concentrations) varied inversely and were linear with exponential growth rate. At high tungstate concentration, when the exponential growth rate approached zero (as a result of negligible 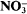 reduction), β
obs approached 1·0197. In the absence of tungstate, β
obs was equal to 1·0037. From this value, we calculated the ratio of efflux to influx to be about 0·19. That is, in the absence of tungstate, more than 80% of the
reduction), β
obs approached 1·0197. In the absence of tungstate, β
obs was equal to 1·0037. From this value, we calculated the ratio of efflux to influx to be about 0·19. That is, in the absence of tungstate, more than 80% of the 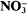 entering the cell underwent reduction with less than 20% released to the medium from the cell. This approach can be used to distinguish changes in influx from changes in metabolic steps in response to factors which affect net uptake rate.
entering the cell underwent reduction with less than 20% released to the medium from the cell. This approach can be used to distinguish changes in influx from changes in metabolic steps in response to factors which affect net uptake rate.
-
- Systematics
-
-
-
The use of 16S ribosomal RNA analyses to investigate the phylogeny of the family Legionellaceae
More LessThe 16S ribosomal RNA sequences of Legionella pneumophila, L. erythra, L. hackeliae, L. spiritensis, L. longbeachae, L. bozemanii (Fluoribacter bozemanae) and L. micdadei (Tatlockia micdadei) were determined using reverse transcriptase. The sequences were compared with published sequences for Gram-negative bacteria and phylogenetic trees were constructed. The data confirm previous work which showed that the family Legionellaceae forms a monophyletic subgroup within the gamma subdivision of the Proteobacteria. The data show that all of the legionellae studied are highly related (> 95 %) on the basis of 16S rRNA sequences and do not support the division of the family Legionellaceae into three genera.
-
-
-
-
Phylogenetic analysis of five medically important Candida species as deduced on the basis of small ribosomal subunit RNA sequences
More LessThe classification of species belonging to the genus Candida Berkhout is problematic. Therefore, we have determined the small ribosomal subunit RNA (srRNA) sequences of the type strains of three human pathogenic Candida species; Candida krusei, C. lusitaniae and C. tropicalis. The srRNA sequences were aligned with published eukaryotic srRNA sequences and evolutionary trees were inferred using a matrix optimization method. An evolutionary tree comprising all available eukaryotic srRNA sequences, including two other pathogenic Candida species, C. albicans and C. glabrata, showed that the yeasts diverge rather late in the course of eukaryote evolution, namely at the same depth as green plants, ciliates and some smaller taxa. The cluster of the higher fungi consists of 10 ascomycetes and ascomycete-like species with the first branches leading to Neurospora crassa, Pneumocystis carinii, Candida lusitaniae and C. krusei, in that order. Next there is a dichotomous divergence leading to a group consisting of Torulaspora delbrueckii, Saccharomyces cerevisiae, C. glabrata and Kluyveromyces lactis and a smaller group comprising C. tropicalis and C. albicans. The divergence pattern obtained on the basis of srRNA sequence data is also compared to various other chemotaxonomic data.
-
Volumes and issues
-
Volume 170 (2024)
-
Volume 169 (2023)
-
Volume 168 (2022)
-
Volume 167 (2021)
-
Volume 166 (2020)
-
Volume 165 (2019)
-
Volume 164 (2018)
-
Volume 163 (2017)
-
Volume 162 (2016)
-
Volume 161 (2015)
-
Volume 160 (2014)
-
Volume 159 (2013)
-
Volume 158 (2012)
-
Volume 157 (2011)
-
Volume 156 (2010)
-
Volume 155 (2009)
-
Volume 154 (2008)
-
Volume 153 (2007)
-
Volume 152 (2006)
-
Volume 151 (2005)
-
Volume 150 (2004)
-
Volume 149 (2003)
-
Volume 148 (2002)
-
Volume 147 (2001)
-
Volume 146 (2000)
-
Volume 145 (1999)
-
Volume 144 (1998)
-
Volume 143 (1997)
-
Volume 142 (1996)
-
Volume 141 (1995)
-
Volume 140 (1994)
-
Volume 139 (1993)
-
Volume 138 (1992)
-
Volume 137 (1991)
-
Volume 136 (1990)
-
Volume 135 (1989)
-
Volume 134 (1988)
-
Volume 133 (1987)
-
Volume 132 (1986)
-
Volume 131 (1985)
-
Volume 130 (1984)
-
Volume 129 (1983)
-
Volume 128 (1982)
-
Volume 127 (1981)
-
Volume 126 (1981)
-
Volume 125 (1981)
-
Volume 124 (1981)
-
Volume 123 (1981)
-
Volume 122 (1981)
-
Volume 121 (1980)
-
Volume 120 (1980)
-
Volume 119 (1980)
-
Volume 118 (1980)
-
Volume 117 (1980)
-
Volume 116 (1980)
-
Volume 115 (1979)
-
Volume 114 (1979)
-
Volume 113 (1979)
-
Volume 112 (1979)
-
Volume 111 (1979)
-
Volume 110 (1979)
-
Volume 109 (1978)
-
Volume 108 (1978)
-
Volume 107 (1978)
-
Volume 106 (1978)
-
Volume 105 (1978)
-
Volume 104 (1978)
-
Volume 103 (1977)
-
Volume 102 (1977)
-
Volume 101 (1977)
-
Volume 100 (1977)
-
Volume 99 (1977)
-
Volume 98 (1977)
-
Volume 97 (1976)
-
Volume 96 (1976)
-
Volume 95 (1976)
-
Volume 94 (1976)
-
Volume 93 (1976)
-
Volume 92 (1976)
-
Volume 91 (1975)
-
Volume 90 (1975)
-
Volume 89 (1975)
-
Volume 88 (1975)
-
Volume 87 (1975)
-
Volume 86 (1975)
-
Volume 85 (1974)
-
Volume 84 (1974)
-
Volume 83 (1974)
-
Volume 82 (1974)
-
Volume 81 (1974)
-
Volume 80 (1974)
-
Volume 79 (1973)
-
Volume 78 (1973)
-
Volume 77 (1973)
-
Volume 76 (1973)
-
Volume 75 (1973)
-
Volume 74 (1973)
-
Volume 73 (1972)
-
Volume 72 (1972)
-
Volume 71 (1972)
-
Volume 70 (1972)
-
Volume 69 (1971)
-
Volume 68 (1971)
-
Volume 67 (1971)
-
Volume 66 (1971)
-
Volume 65 (1971)
-
Volume 64 (1970)
-
Volume 63 (1970)
-
Volume 62 (1970)
-
Volume 61 (1970)
-
Volume 60 (1970)
-
Volume 59 (1969)
-
Volume 58 (1969)
-
Volume 57 (1969)
-
Volume 56 (1969)
-
Volume 55 (1969)
-
Volume 54 (1968)
-
Volume 53 (1968)
-
Volume 52 (1968)
-
Volume 51 (1968)
-
Volume 50 (1968)
-
Volume 49 (1967)
-
Volume 48 (1967)
-
Volume 47 (1967)
-
Volume 46 (1967)
-
Volume 45 (1966)
-
Volume 44 (1966)
-
Volume 43 (1966)
-
Volume 42 (1966)
-
Volume 41 (1965)
-
Volume 40 (1965)
-
Volume 39 (1965)
-
Volume 38 (1965)
-
Volume 37 (1964)
-
Volume 36 (1964)
-
Volume 35 (1964)
-
Volume 34 (1964)
-
Volume 33 (1963)
-
Volume 32 (1963)
-
Volume 31 (1963)
-
Volume 30 (1963)
-
Volume 29 (1962)
-
Volume 28 (1962)
-
Volume 27 (1962)
-
Volume 26 (1961)
-
Volume 25 (1961)
-
Volume 24 (1961)
-
Volume 23 (1960)
-
Volume 22 (1960)
-
Volume 21 (1959)
-
Volume 20 (1959)
-
Volume 19 (1958)
-
Volume 18 (1958)
-
Volume 17 (1957)
-
Volume 16 (1957)
-
Volume 15 (1956)
-
Volume 14 (1956)
-
Volume 13 (1955)
-
Volume 12 (1955)
-
Volume 11 (1954)
-
Volume 10 (1954)
-
Volume 9 (1953)
-
Volume 8 (1953)
-
Volume 7 (1952)
-
Volume 6 (1952)
-
Volume 5 (1951)
-
Volume 4 (1950)
-
Volume 3 (1949)
-
Volume 2 (1948)
-
Volume 1 (1947)
Most Read This Month
