 -
Volume 137,
Issue 5,
1991
-
Volume 137,
Issue 5,
1991

Volume 137, Issue 5, 1991
- Review Article
-
- Biochemistry
-
-
-
Methylation of phenol by chloromethane in the fungus Phellinus pomaceus
More LessThe kinetics of phenol methylation by chloromethane (CH3Cl) in intact mycelia of the fungus Phellinus pomaceus were examined. Anisole was produced linearly with respect to time at a rate of 6·3 nmol g−1 h−1 over 16 h when washed mycelia were incubated with phenol and C2H3Cl. Incorporation of C2H3- label into anisole attained a plateau value of 27% within 2 h. The rate of anisole production under N2 was only 36% of that in air, and the proportion of C2H3- label incorporated from exogenous C2H3- increased from 30 to 45% under these conditions. The rate of methylation attained a maximum in 10 mm-phenol, but the level of activity was only about 10% of that of the fungal carboxyl-methylating system. The presence of exogenous C2H3Cl clearly stimulated anisole formation, demonstrating that the rate of CH3Cl biosynthesis limited methylation to some extent. In contrast to the carboxyl-methylating system, no significant inhibition of methylation was observed at methyl acceptor concentrations up to 17·5 mm, and no sharp fall in the rate of gaseous C2H3Cl release or rapid increase in C2H3-incorporation were observed at supraoptimal concentrations of acceptor, indicating that endogenous CH3Cl biosynthesis was not inhibited under these conditions. Investigations of the rate of C2H3- incorporation into anisole from exogenous C2H3Cl showed a linear relationship between the logarithm of % C2H3- incorporation and the logarithm of C2H3Cl concentration, suggesting that, as postulated for the carboxyl-methylating system, the CH3Cl-synthesizing and CH3Cl-utilizing enzyme systems were situated on either side of a membrane within the cell.
-
-
-
-
Cell wall glycoproteins of Candida albicans as released by different methods
More LessDifferent methods of extraction frequently used in other studies were used to release glycoproteins from both intact cells and isolated cell walls of yeast and hyphal forms of Candida albicans. Extracts were obtained from whole cells by treatment (i) with 2-mercaptoethanol (βME) at pH 8.6 and 37 °C and (ii) with zymolyase after treatment with βME. Extracts were obtained from isolated and washed cell walls (i) by boiling with βME and sodium dodecyl sulphate (SDS), (ii) by boiling with SDS and (iii) by treatment with zymolyase after SDS. The extracts were separated by SDS-polyacrylamide gel electrophoresis and analysed by Western blotting with four reagents. Analysis with concanavalin A (ConA) revealed different glycoprotein populations depending on the treatment. Three possible germ-tube-specific constituents were observed; an 80 kDa component released by βME from both intact cells and cell walls, and 47 kDa and 43 kDa moieties released by zymolyase only from intact cells. MAb 4C12, specific for the protein portion of a large germ tube constituent, recognized polydisperse material which just entered the gel in βME extracts and in the region extending up from 200 kDa to near the top of the gel in zymolyase extracts. MAb 24.17, specific for a carbohydrate determinant of yeast phase cells, reacted with disperse material in the region from the top of the gel to one-third to two-thirds the distance to the 220 kDa mass marker. Antiserum specific for the serotype A determinant of mannan reacted with large disperse component(s) migrating in the region from the top of the gel to about two-thirds the distance to the 220 kDa mass marker and with a 180 kDa component. The components recognized by MAb 4C12, but not those recognised by MAb 24.17 and serotype A antiserum, were effected by treatment with endo-β-N-acetylglucosamidase H. The various analyses revealed that the method of extraction affected the composition and size of the constituents recognized by the reagents.
-
-
-
Heterogeneity of proteinases from the hyperthermophilic archaeobacterium Pyrococcus furiosus
More LessIntracellular and extracellular samples from the extremely thermophilic archaeobacterium Pyrococcus furiosus showed the presence of multiple active proteinases. Using gelatin-containing SDS-PAGE, up to 13 activity bands were visualized with apparent molecular masses of between 66 and 135 kDa. Characterization studies revealed these bands to be due to discrete polypeptides, and not artefacts. Results from gel permeation chromatography, sucrose density gradient centrifugation and non-denaturing PAGE suggested that some of these proteohtic polypeptides may exist as active aggregates either in vivo or in vitro before being dissociated by SDS to active monomers.
-
-
-
Pathways of glutamate catabolism among Fusobacterium species
More LessGlutamate is a major source of energy for Fusobacterium species but its mode of catabolism has not hitherto been elucidated. Cell suspensions of F. nucleatum and F. varium, as representative species from the oral cavity and gastrointestinal tract, respectively, both decarboxylated position-labelled glutamate but by different pathways. 14CO2 was released only from C-5 by F. nucleatum whereas F. varium decarboxylated glutamate at either C-1 or C-5. In both species, 2 mols of glutamate fermented yielded 2 mols of acetate and 1 mol of butyrate, suggesting the possibility of three metabolic pathways: the 2-oxoglutarate, mesaconate and 4-aminobutyrate pathways. Enzymes representative of the three pathways were assayed for in cell-free extracts of fusobacteria. All species tested possessed high levels of both glutamate dehydrogenase and 2-oxoglutarate reductase, indicating the presence of the 2-oxoglutarate pathway. Enzymes representative of the mesaconate pathway were detected in F. sulci, F. ulcerans, F. mortiferum and F. varium, while the latter two species also possessed the 4-aminobutyrate pathway. The pathways of glutamate catabolism therefore bore no relationship to the site of isolation of the fusobacteria tested but instead correlated with their chemotaxonomic properties. Thus, F. varium, F. mortiferum, F. ulcerans and F. sulci, which possess a peptidoglycan structure based on diaminopimelic acid, have either two or three pathways for glutamate catabolism whereas F. nucleatum and other species that have a lanthionine-based murein metabolized glutamate solely by the 2-oxoglutarate pathway.
-
-
-
Purification and characterization of aminopeptidase A from Lactococcus lactis subsp. lactis NCDO 712
More LessAn aminopeptidase A (EC 3.4.11.7) was purified from Lactococcus lactis subsp. lactis NCDO 712. Of the 18 aminoacyl-alanine dipeptides tested, the enzyme hydrolysed Asp-Ala, Glu-Ala and Ser-Ala. It was also active against tripeptide substrates but did not hydrolyse Ala-Asp or Ala-Glu. The kinetics of dipeptide hydrolysis were allosteric with positive cooperativity of substrate binding. The Hill constants were 1.52 for Asp-Ala, 1.51 for Glu-Ala and 1.61 for Ser-Ala. The M r was found to be 245000 by gel-filtration chromatography. A single band corresponding to an M r of 41 000 was detected by SDS-PAGE of the purified enzyme which indicated that the enzyme is hexameric. Activity against glutamate p-nitroanilide was optimum at 65 °C and pH 8. Activity was inhibited by 1 mM-EDTA, implying that the enzyme is metal-containing. Cu2+, Mn2+ and Zn2+ (all at 1 mM) inhibited activity and Co2+ was stimulatory.
-
- Biotechnology
-
-
-
Isolation and purification of viable Ureaplasma urealyticum cells free from medium components
More LessA procedure was devised to produce Ureaplasma urealyticum preparations free of adsorbed components of the growth medium, which contains high concentrations of serum. The ureaplasmas were cultivated in a medium containing PPLO-serum fraction as a replacement for horse serum. High titres of ureaplasmas (> 107 c.f.u. ml-1) were obtained. Harvested cells were then purified by Urografin density gradient centrifugation. By use of 3H-labelled ureaplasma cells and 125I-labelled medium components, a distinct band of viable cells devoid of serum constituents was demonstrated. The absence of medium components was verified by immunoblotting cells from this band with antiserum to medium components. Medium components that had been present before the purification procedure were undetectable in the purified cell fraction obtained. The viability of the purified ureaplasma cells represented an 85% recovery rate and their antigenicity, examined with anti-serotype specific antiserum, remained intact. This easy and reproducible procedure can be used to prepare purified ureaplasmas for investigation of ureaplasmal antigens and their expression and/or role in disease.
-
-
-
-
One-step purification of cholesterol oxidase from culture broth of a Pseudomonas sp. using a novel affinity chromatography method
More LessCholesterol oxidase from the culture broth of a Pseudomonas sp. was purified with a yield of more than 70% by a one-step procedure using a column of cholesterylglycine-carboxymethylcellulose; active enzyme was eluted by Triton X-100. The purified enzyme was homogeneous by SDS-PAGE.
-
- Development And Structure
-
-
-
Induction of sclerotia in Sclerotium rolfsii by short low-temperature treatment
More LessWhen grown on two agar media in Petri dishes a Sclerotium rolfsii Sacc. isolate produced sclerotia in different patterns, predominantly at the colony centre on one medium (glucose/yeast extract) but at the dish wall on the other (glucose/ammonium nitrate). However, on both media a short cold treatment (3 h at 5 °C) of colonies grown at 24 °C temporarily interrupted growth and induced subsequent sclerotial formation in a ring where the colony margin was at the time of the cold shock. A second cold treatment 1 d after the first induced a sclerotial ring but negated the effect of the first treatment. The results support the view that interruption of hyphal growth triggers sclerotial formation.
-
-
-
-
Tubular spinae are long-distance connectors between bacteria
More LessThe marine pseudomonad D71 (NCMB 2018) [‘Spinomonas maritima’] can be induced to produce long tubular surface appendages (spinae) in a growth medium of low osmolarity. In general, spina-carrying cells show these appendages with open distal ends. We examined cultured cells by scanning and transmission electron microscopy, using both critical-point drying and thin sectioning after embedding with agarose protection. By scanning electron microscopy, spinae were observed that connected cells over distances of several micrometers. Ultrathin sections often revealed an additional layer outside the outer membrane, resembling an S-layer. The inner and outer cell membranes were often joined at spina-insertion areas. Furthermore, evidence was found in ultrathin sections for uninterrupted tubes connecting two cells over a distance of up to 7 μm. We propose, therefore, that spinae form the framework for wide open cell clusters; we hypothesize that these spinae might also permit an exchange of cell-to-cell signals.
-
- Genetics And Molecular Biology
-
-
-
Isolation and evolutionary analysis of a RepFVIB replicon of the plasmid pSU212
More LessWe have isolated at least two different replication regions from pSU401, a Tn802 insertion derivative of the IncFVI plasmid pSU212. One of the replication regions (RepFVIB) is highly homologous to the RepFIIA replicon of IncFII plasmids, and thus belongs to the RepFIIA family. We have also cloned the incompatibility determinant (incFVI) and the copy number control gene (cop) from RepFVIB and determined their nucleotide sequences. The analysis of the sequences supports the idea of a modular evolution of RepFIIA plasmids.
-
-
-
-
Nucleotide sequence of a repetitive element isolated from Leptospira interrogans serovar hardjo type hardjo-bovis
More LessA repetitive element from the genome of Leptospira interrogans serovar hardjo type hardjo-bovis (‘L. hardjo-bovis’) was identified, cloned and sequenced. Similar sequences were shown by hybridization to be encoded by a further eight of 32 other leptospiral serovars tested. An undefined number of repetitive elements were located in the L. hardjo-bovis genome; sequence degeneracy of the elements was observed and no significant open reading frames were identified within the AT-rich (60%) 1467 bp repetitive element. The termini encoded a GC-rich 8 bp repeat motif and two variants showed rearrangements centred on these motifs. The nucleotide sequences of the chromosomal regions flanking the repetitive elements were determined but showed no similarities, with one exception which had a GAAC repeat directly adjacent to both termini. Similar hybridization patterns were shown by Southern transfers of L. hardjo-bovis total genomic digests probed with the repetitive element. Oligonucleotide primer pairs designed from sequences internal to the repetitive element and adjacent chromosomal regions were used in polymerase chain reaction experiments. With one primer pair all L. hardjo-bovis isolates, but no other serovar, gave identical amplified products. Evidence that the repetitive element may have derived from an acquired insertion sequence that is now inactive and chromosomally fixed is discussed.
-
-
-
Chromosomal insertion of TOL transposons in Pseudon eruginosa PAO
More LessInsertions of the TOL plasmid transposons Tn4651 and Tn4653 into the Pseudomonas aeruginosa PAO chromosome were isolated by a temperature selection technique. The locations and orientations of 16 insertions were determined by pulsed field gel electrophoresis and Southern hybridization with genomic and TOL DNA probes. All insertions occurred within a 334 kb region of the chromosome (representing less than 6% of the genome) with nine of the inserts clustered within a 10 kb area. Each transposon was able to insert in either orientation. An internal duplication of the 39 kb excisable region of pWW0 was seen in two independent insertions.
-
-
-
Regulation of the oxidative stress response by the hpr gene in Bacillus subtilis
More LessBacillus subtilis mutants with null mutations in the spo0A gene are resistant to oxidative stress during the exponential phase of growth. This resistance phenotype can be suppressed by mutations in the abrB gene, or in the hpr gene. Both of these gene products are negative regulatory proteins which are over-produced in a spo0A strain, and the over-production of the hpr gene product results from over-production of the abrB gene product. The results suggested that the resistance to oxidative stress in a spo0A strain is due to the lack of a protein directly controlled by the hpr negative regulator. Other mutations in the spo0A gene conferring resistance to ethanol stress (eth) or suppressors of sporulation phenotypes (sof) had no effect on the sensitivity to oxidative stress of strains bearing them.
-
-
-
Bacillus licheniformis APase I gene promoter: a strong well-regulated promoter in B. subtilis
More LessThe 5' regulatory region and the portion of the structural gene coding for the amino-terminal sequence of alkaline phosphatase I (APase I) were isolated from Bacillus licheniformis MC14 using a synthetic oligodeoxynucleotide deduced from the amino acid sequence of the enzyme. The DNA sequence analysis of this region revealed an open reading frame of 129 amino acids containing the amino-terminal sequence of the mature APase protein. The protein sequence was preceded by a putative signal sequence of 32 amino acid residues. The predicted amino acid sequence of the partial APase clone as well as the experimentally determined amino acid sequence of the enzyme indicated that B. licheniformis APase retains the important features conserved among other APases of Bacillus subtilis, Escherichia colil Saccharomyces cerevisiae, and various human tissues. Heterologous expression studies of the promoter using a fusion with the lacZ gene indicated that it functions as a very strong inducible promoter in B. subtilis that is tightly regulated by phosphate concentration.
-
-
-
Identification of aecA mutations in Bacillus subtilis as nucleotide substitutions in the untranslated leader region of the aspartokinase II operon
More LessRecent genetic mapping of the aspartokinase II (lysC) operon of Bacillus subtilis [M. Petricek. L. Rutberg & L. Hederstedt (1989) FEMS Microbiology Letters 61,85–88; N. Y. Chen. J. J. Zhang & H. Paulus (1989) Journal of General Microbiology 135, 2931–2940] has shown its chromosomal location to be close to the aecA locus, the mutation of which leads to highly increased levels of aspartokinase II. In order to examine the relationship between lysC and aecA, we have cloned the control regions of the lysC operon from several independent aecA mutants and determined their nucleotide sequences. The nucleotide sequences of the aecA mutants differed from the wild-type sequence by the substitution of one or two nucleotides at two widely separated sites in the transcribed leader region of the lysC operon. To confirm that the observed nucleotide changes are indeed responsible for the AecA phenotype and not simply the reflection of sequence polymorphisms in different B. subtilis strains, we introduced the same nucleotide substitutions as those observed in the aecA strains into the leader region of the wild-type lysC operon by oligonucleotide-directed mutagenesis. The expression of the mutagenized genes was analysed after transcriptional or translational fusion to lacZ in a single-copy integration vector. The levels of β-galactosidase were greatly elevated by the nucleotide substitutions, with similar increases observed in transcriptional and translational fusions. The high level of expression of β-galactosidase in the lysC-lac’Z strains with nucleotide substitutions corresponding to the aecA mutations was resistant to repression by L-lysine but was completely abolished by the inactivation of the lysC promoter. These results suggest that the sites identified by nucleotide sequencing of the aecA mutants are important regulatory sites in the lysC operon, which exert their effect at the level of transcription involving the normal lysC promoter.
-
- Pathogenicity And Medical Microbiology
-
-
-
Ultrastructural localization of the Pasteurella multocida toxin in a toxin-producing strain
More LessToxigenic strains of Pasteurella multocida produce the 147 kDa protein Pasteurella multocida toxin (PMT) which is responsible for the osteoclastic bone resorption in progressive atrophic rhinitis in pigs and induces such resorption in all experimental animals tested so far. In the present study we have carried out immunocytochemistry on formaldehyde- and glutaraldehyde-fixed ultracryocut P. multocida using a pool of monoclonal antibodies against different epitopes on PMT as the first layer and affinity purified rabbit anti-mouse IgG as the second layer. Goat anti-rabbit IgG conjugated with 5 nm gold particles was used as marker. The gold particles were silver-enhanced prior to examination in the transmission electron microscope. Whole bacteria were also immunostained after fixation and critical point drying and examined by scanning transmission electron microscopy. The results showed that PMT was located in the cytoplasm of P. multocida. PMT could not be detected on intact, undamaged P. multocida by scanning electron microscopy. Neither pili nor flagella could be detected on the surface of the negatively stained P. multocida strains investigated. PMT has a series of characteristics encompassed in the definition of an exotoxin. However, that PMT was not secreted by living intact P. multocida is unexpected for an exotoxin.
-
-
-
-
Siderophore production by Aeromonas salmonicida
More LessGrowth under conditions of iron-restriction and the production of siderophores was examined in 21 typical and 14 atypical strains of Aeromonas salmonicida. With the exception of one atypical strain, all strains grew and multiplied in the presence of the high-affinity iron chelators ethylenediamine di(o-hydroxyphenylaectic acid), α,α’-dipyridyl or transferrin. Chrome azurol S agar was used to screen bacterial strains growing under these conditions for the production of siderophores. Siderophore production was detected only in the typical strains. Siderophores were also detected in the iron-restricted culture supernatants of typical strains, where they were associated with an iron-binding activity. The siderophore was extracted from iron-restricted culture supernatant of one strain by adsorption onto an XAD-7 resin; it behaved as a 2,3-diphenol-catechol in several colorimetric assays. The results indicate that although both typical and atypical strains of A. salmonicida grow and multiply under conditions of iron-restriction, they use different iron-uptake mechanisms, siderophore-mediated and siderophore-independent, respectively. In cross-feeding assays, growth of typical strains was stimulated only by homologous iron-restricted supernatant, suggesting strain differences in the siderophore produced. However, one strain produced a culture supernatant with growth-stimulating activity for other typical and also atypical strains.
-
- Physiology And Growth
-
-
-
Chemiluminescence and superoxide production in Acanthamoeba castellanii: free radicals generated during oxidative stress
More LessThe amoeba Acanthamoeba castellanii generated both luminol- and lucigenin-enhanced chemiluminescence upon addition of the respiratory inhibitor sodium cyanide, but not upon the addition of sodium azide. Photon emission in the presence of lucigenin was three- to fourfold greater than that measured in the presence of luminol, but both forms of chemiluminescence were strictly dependent upon the presence of O2, indicating the requirement for oxidative reactions in these processes. Lucigenin-chemilununescence measured during the phagocytosis of latex beads under identical conditions was, however, barely detectable above background levels. Cyanide similarly induced the formation of
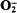 , as indicated by the stimulation of superoxide-dismutase-inhibitable cytochrome c reduction, and the rate of production was similar to that previously observed during phagocytosis of latex particles or yeasts by these cells. Thus, in view of the similar rates of
, as indicated by the stimulation of superoxide-dismutase-inhibitable cytochrome c reduction, and the rate of production was similar to that previously observed during phagocytosis of latex particles or yeasts by these cells. Thus, in view of the similar rates of 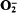 production during cyanide exposure or phagocytosis, but disparate rates of lucigenin-chemiluminescence, these two treatments must activate different molecular processes leading to reactive oxidant production. The rates of cyanide-stimulated lucigenin-chemiluminescence were directly proportional to the O2 tensions in the medium from 0 to 300 μM, indicating that the rates of free-radical-generating reactions were directly related to the oxygen tensions in the environment. The superoxide dismutase inhibitor diethyldithiocarbamate similarly stimulated lucigenin-chemiluminescence, with photon emission again being dependent upon O2 tensions in the range 0–320 μM. A mechanism by which cells may limit
production during cyanide exposure or phagocytosis, but disparate rates of lucigenin-chemiluminescence, these two treatments must activate different molecular processes leading to reactive oxidant production. The rates of cyanide-stimulated lucigenin-chemiluminescence were directly proportional to the O2 tensions in the medium from 0 to 300 μM, indicating that the rates of free-radical-generating reactions were directly related to the oxygen tensions in the environment. The superoxide dismutase inhibitor diethyldithiocarbamate similarly stimulated lucigenin-chemiluminescence, with photon emission again being dependent upon O2 tensions in the range 0–320 μM. A mechanism by which cells may limit 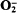 generating reactions, and so reduce damaging free radical reactions, was observed when anaerobic suspensions were reaerated. These data indicate that oxidative stress and phagocytosis provide two intracellular sources of free-radical generating reactions in these cells.
generating reactions, and so reduce damaging free radical reactions, was observed when anaerobic suspensions were reaerated. These data indicate that oxidative stress and phagocytosis provide two intracellular sources of free-radical generating reactions in these cells.
-
-
-
-
13C NMR analysis of a developmental pathway mutation in Saccharomyces cerevisiae reveals a cell derepressed for succinate dehydrogenase
More Less13C nuclear magnetic resonance (NMR) spectroscopy was used to study the metabolism of [2-13C]acetate in a diploid strain of Saccharomyces cerevisiae homozygous for the spo50 mutation. This mutation results in failure to initiate sporulation and suppresses spd mutations (which cause derepressed sporulation). By analysing the pattern of 13C-labelling in glutamate it was deduced that the glyoxylate cycle is responsible for most of the acetate utilization and that there is very little tricarboxylic acid cycle activity. The labelling of α,α′-trehalose indicated that gluconeogenesis and the hexose monophosphate pathway operate in a similar way to the wild-type. The mutant strain has higher levels of succinate dehydrogenase than the wild-type. All of the physiological alterations caused by the spo50 mutation can be explained by this difference.
-
Volumes and issues
-
Volume 170 (2024)
-
Volume 169 (2023)
-
Volume 168 (2022)
-
Volume 167 (2021)
-
Volume 166 (2020)
-
Volume 165 (2019)
-
Volume 164 (2018)
-
Volume 163 (2017)
-
Volume 162 (2016)
-
Volume 161 (2015)
-
Volume 160 (2014)
-
Volume 159 (2013)
-
Volume 158 (2012)
-
Volume 157 (2011)
-
Volume 156 (2010)
-
Volume 155 (2009)
-
Volume 154 (2008)
-
Volume 153 (2007)
-
Volume 152 (2006)
-
Volume 151 (2005)
-
Volume 150 (2004)
-
Volume 149 (2003)
-
Volume 148 (2002)
-
Volume 147 (2001)
-
Volume 146 (2000)
-
Volume 145 (1999)
-
Volume 144 (1998)
-
Volume 143 (1997)
-
Volume 142 (1996)
-
Volume 141 (1995)
-
Volume 140 (1994)
-
Volume 139 (1993)
-
Volume 138 (1992)
-
Volume 137 (1991)
-
Volume 136 (1990)
-
Volume 135 (1989)
-
Volume 134 (1988)
-
Volume 133 (1987)
-
Volume 132 (1986)
-
Volume 131 (1985)
-
Volume 130 (1984)
-
Volume 129 (1983)
-
Volume 128 (1982)
-
Volume 127 (1981)
-
Volume 126 (1981)
-
Volume 125 (1981)
-
Volume 124 (1981)
-
Volume 123 (1981)
-
Volume 122 (1981)
-
Volume 121 (1980)
-
Volume 120 (1980)
-
Volume 119 (1980)
-
Volume 118 (1980)
-
Volume 117 (1980)
-
Volume 116 (1980)
-
Volume 115 (1979)
-
Volume 114 (1979)
-
Volume 113 (1979)
-
Volume 112 (1979)
-
Volume 111 (1979)
-
Volume 110 (1979)
-
Volume 109 (1978)
-
Volume 108 (1978)
-
Volume 107 (1978)
-
Volume 106 (1978)
-
Volume 105 (1978)
-
Volume 104 (1978)
-
Volume 103 (1977)
-
Volume 102 (1977)
-
Volume 101 (1977)
-
Volume 100 (1977)
-
Volume 99 (1977)
-
Volume 98 (1977)
-
Volume 97 (1976)
-
Volume 96 (1976)
-
Volume 95 (1976)
-
Volume 94 (1976)
-
Volume 93 (1976)
-
Volume 92 (1976)
-
Volume 91 (1975)
-
Volume 90 (1975)
-
Volume 89 (1975)
-
Volume 88 (1975)
-
Volume 87 (1975)
-
Volume 86 (1975)
-
Volume 85 (1974)
-
Volume 84 (1974)
-
Volume 83 (1974)
-
Volume 82 (1974)
-
Volume 81 (1974)
-
Volume 80 (1974)
-
Volume 79 (1973)
-
Volume 78 (1973)
-
Volume 77 (1973)
-
Volume 76 (1973)
-
Volume 75 (1973)
-
Volume 74 (1973)
-
Volume 73 (1972)
-
Volume 72 (1972)
-
Volume 71 (1972)
-
Volume 70 (1972)
-
Volume 69 (1971)
-
Volume 68 (1971)
-
Volume 67 (1971)
-
Volume 66 (1971)
-
Volume 65 (1971)
-
Volume 64 (1970)
-
Volume 63 (1970)
-
Volume 62 (1970)
-
Volume 61 (1970)
-
Volume 60 (1970)
-
Volume 59 (1969)
-
Volume 58 (1969)
-
Volume 57 (1969)
-
Volume 56 (1969)
-
Volume 55 (1969)
-
Volume 54 (1968)
-
Volume 53 (1968)
-
Volume 52 (1968)
-
Volume 51 (1968)
-
Volume 50 (1968)
-
Volume 49 (1967)
-
Volume 48 (1967)
-
Volume 47 (1967)
-
Volume 46 (1967)
-
Volume 45 (1966)
-
Volume 44 (1966)
-
Volume 43 (1966)
-
Volume 42 (1966)
-
Volume 41 (1965)
-
Volume 40 (1965)
-
Volume 39 (1965)
-
Volume 38 (1965)
-
Volume 37 (1964)
-
Volume 36 (1964)
-
Volume 35 (1964)
-
Volume 34 (1964)
-
Volume 33 (1963)
-
Volume 32 (1963)
-
Volume 31 (1963)
-
Volume 30 (1963)
-
Volume 29 (1962)
-
Volume 28 (1962)
-
Volume 27 (1962)
-
Volume 26 (1961)
-
Volume 25 (1961)
-
Volume 24 (1961)
-
Volume 23 (1960)
-
Volume 22 (1960)
-
Volume 21 (1959)
-
Volume 20 (1959)
-
Volume 19 (1958)
-
Volume 18 (1958)
-
Volume 17 (1957)
-
Volume 16 (1957)
-
Volume 15 (1956)
-
Volume 14 (1956)
-
Volume 13 (1955)
-
Volume 12 (1955)
-
Volume 11 (1954)
-
Volume 10 (1954)
-
Volume 9 (1953)
-
Volume 8 (1953)
-
Volume 7 (1952)
-
Volume 6 (1952)
-
Volume 5 (1951)
-
Volume 4 (1950)
-
Volume 3 (1949)
-
Volume 2 (1948)
-
Volume 1 (1947)
Most Read This Month
