 -
Volume 131,
Issue 3,
1985
-
Volume 131,
Issue 3,
1985

Volume 131, Issue 3, 1985
- Physiology And Growth
-
-
-
The all2 Gene is Required for the Induction of the Purine Deamination Pathway in Schizosaccharomyces pombe
More LessFive mutants were isolated at the all2 gene on the basis of their inability to utilize hypoxanthine as a sole source of nitrogen. These mutants failed to utilize the purines adenine, hypoxanthine, xanthine, uric acid, allantoin and allantoic acid, although they could utilize urea and ammonium. The all2 mutants appeared to be defective in purine induction of uricase, allantoinase, allantoicase and ureidoglycollase activities but retained wild-type activity of the constitutively synthesized urease. The all2 mutations were recessive.
-
-
-
-
Competence-specific Autolysis in Streptococcus sanguis
More LessStreptococcus sanguis strain Wicky activated to competence for genetic transformation is known to undergo a rapid decrease in optical density upon transfer to an alkaline buffer containing reducing agents. We studied the mechanism of this autolysis-like process and made the following observations. The process was specific because preincubation of the competence inducing factor with a specific inactivating protein prevented both cellular lysis and acquisition of competence for genetic transformation. The optical density decrease of competent bacteria involved the release of a large fraction of intracellular protein, RNA and lipid. However, no hydrolysis of phospholipid and no degradation of cell wall polymers including peptidoglycan could be detected. No peptidoglycan hydrolase activity capable of degrading radiolabeled S. sanguis cell walls was detected in unfractionated S. sanguis extracts. It is suggested that autolysis of competent S. sanguis involves the activity of a novel type of murein hydrolase that introduces only a limited number of bond breaks into the peptidoglycan.
-
-
-
Characterization of DNA Fragments Encoding Fimbriae of the Uropathogenic Escherichia coli Strain KS71
More LessRecombinant plasmids were constructed that expressed the KS71A, KS71B and KS71C fimbrial antigens of the pyelonephritogenic Escherichia coli strain KS71 (04:K12) in E. coli HB101. The KS71C-encoding genes were located on a 6·4 kb HindIII–XhoI fragment obtained from the recombinant cosmid pKTH145 that expresses this antigen. Spontaneous KS71C– mutants were isolated that contained a 0·8 kb insert in a specific restriction fragment of KS71C-encoding recombinant plasmids. The KS71 B-encoding segment was located on a 11·5 kb deletable DNA fragment of recombinant cosmid pKTH144. A DNA fragment encoding the KS71A fimbria was obtained on a 12 kb EcoRI fragment of the recombinant cosmid expressing this antigen in E. coli HB101 and closely resembled the KS71B-encoding fragment. In the recombinant cosmid, the KS71 B-expressing region was flanked by homologous DNA segments. A similar stretch of DNA was found close to the KS71A-expressing DNA region.
-
-
-
Isolation and Characterization of Small Heat-stable Acid-soluble DNA-binding Proteins from Bacillus subtilis Nucleoids
More LessSmall heat-stable, acid-soluble proteins (HASP) have been isolated from Bacillus subtilis nucleoids obtained from cell lysates of low ionic strength and lysozyme concentration. They were identified by their ability to bind homologous and heterologous native and denatured DNA. Four major species, of 8·5, 12, 23 and 26 kDal, were found. Their affinity for DNA was moderate as measured by the sensitivity to ionic strength of the DNA-protein complex (0·1-0·4-m-NaCl). Partial digestion by micrococcal nuclease of the ‘low ionic strength nucleoids’ released a DNA fragment of 80-120 bp. The data reported here indicate that small basic proteins, together with other components such as RNA, cations and polyamines, may be involved in the compaction of the prokaryotic genome.
-
-
-
The Effect of Cerulenin on the Morphogenesis and Autolytic Activity of Bacillus subtilis
More LessPartial inhibition of lipid synthesis in Bacillus subtilis by the inclusion of cerulenin in growth media led to the formation of chains of spheroidal cells both of wild-type strains and lyt mutants. Washed cell suspensions prepared from wild-type cultures treated with cerulenin lysed only very slowly compared with those from control cultures although the activity of autolysins in 5 m-LiCl extracts made from such organisms was only about 30% less than those in extracts from controls. The behaviour of the cultures, such as the separation of cells and the reaction to β-lactams, was as if they were grossly deficient in autolytic activity. The concentration of cerulenin used (7·5 μg ml–1) reduced the growth rate two- to threefold but exponential growth at this slower rate continued for at least 18 h. The steady state concentration of total protein and peptidoglycan per unit bacterial mass was the same as in control cultures but the phospholipid content was reduced by 50%.
-
-
-
Morphological Alterations of the Fission Yeast Schizosaccharomyces pombe in the Presence of Aculeacin A: Spherical Wall Formation
More LessThe fission yeast Schizosaccharomyces pombe, grown in the presence of aculeacin A (Acu), an antifungal antibiotic, forms spherical rather than cylindrical walls. This aberrant morphogenesis was studied in the presence of Acu at 1 μg ml–1 under conditions in which over 90% of the cells continued to grow but with altered morphology. Microscopic observations and chemical analyses revealed that the spherical walls had a looser structure than the cylindrical wall and that their syntheses of alkali-insoluble glucan and mannan were reduced whereas the synthesis of alkali-soluble glucan was enhanced. Spherical walls of glutaraldehyde-fixed cells were hardly digested by Zymolyase 60000 [a (1→3)-β-glucanase] but were digested by Novozym 234 [a (1→3)-α-glucanase]. From these results the components and morphology of the spherical walls are discussed in comparison with the cylindrical walls.
-
-
-
Cytoplasmic Ca2+Homeostasis Maintained by a Vacuolar Ca2+Transport System in the Yeast Saccharomyces cerevisiae
More LessDifferential extraction of Ca2+ from the cytoplasmic and vacuolar pools of the yeast Saccharomyces cerevisiae, using DEAE-dextran, revealed that most of the cellular Ca2+ was bound, precipitated or sequestered within the vacuole. When the concentration of Ca2+ in the medium was raised from 10–6 m to 10–3 m, cytoplasmic Ca2+ homeostasis was maintained at 5·8 × 10–6 to 2·3 × 10–5 m, whereas the vacuoles accumulated higher concentrations of Ca2+. The results indicate that the vacuoles function as a cytoplasmic Ca2+ buffering system and as the major sequestering organelle for Ca2+. A respiratory-deficient mutant (ρ°) displayed a similar intracellular distribution of Ca2+ to the wild-type. When cells were permeabilized by DEAE-dextran the vacuoles were still capable of Ca2+ uptake. This uptake proceeded without the addition of ATP or glucose in fresh preparations but required the addition of ATP after incubation of the permeabilized cells in buffered sorbitol for 2 h. The results are consistent with the proposed Ca2+/H+ antiport in the vacuolar membrane, which is driven by
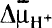 formed by the H+-ATPase pumping H+ into the vacuole.
formed by the H+-ATPase pumping H+ into the vacuole.
-
-
-
The Effect of Pectin and Related Compounds on Encystment and Germination of Phytophthora palmivora Zoospores
More LessCitrus pectin and other uronic acids (polygalacturonate, alginate and galacturonate) accelerated encystment and germination in suspensions of Phytophthora palmivora zoospores. Other anions, polyanions and polysaccharides containing α(1→4) linkages were not effective in this respect. The uronic acids tested differed from one another both in the concentrations required for maximal rates of encystment and in the extent to which they stimulated germination as well as encystment. Pectin-accelerated encystment did not require free Ca2+in the suspending medium, but when the Ca2+concentration exceeded 100 μm, uronate-stimulated encystment and germination was suppressed. This blocking of uronate-accelerated encystment was almost specific for Ca2+, although Sr2+and Ba2+showed some effect. It is proposed that the action of uronates in accelerating encystment of P. palmivora zoospores is analogous to agonists which induce stimulus-mediated secretion in many animal cell types.
-
-
-
Postponement of Cell Division by Nutritional Shift-up in Escherichia coli
More LessNutritional shifts up of synchronized or random populations of Escherichia coli (strain ML30 and derivatives of B/r and K12) to a richer medium were followed (nutritional pulse) or not (shift-up) by shifts down to the original poor medium. In most cases, the first postshift division was clearly postponed by a shift-up or a nutritional pulse. This delay of the first division was almost constant, whatever the cell age at the time of the transition, and, for the nutritional pulse only, whatever the time spent by cells in the rich medium (between 2 and 25% of the generation time characteristic of poor medium). Following a shift-up, the new steady state of division rhythm and of mean cell mass was reached at about the second postshift division, whereas after a nutritional pulse, it took three generations to return to the steady state prevailing in poor medium. When both the poor and the rich medium were varied, the extent of the postponement of cell division after a nutritional pulse increased when the amplitude of the stimulus (i.e. the difference in richness) was increased. This was not the case with a simple shift-up, where it seemed that the postponement was compensated in part by the accelerating effects of the rich medium.
-
- Systematics
-
-
-
The Phylogenetic Position of Streptococcus and Enterococcus
More LessStreptococcus pyogenes, S. equinus, S. bovis, S. salivarius, S. sanguis, S. mutans, S. rattus, S. cricetus, S. lactis, S. raffinolactis and Enterococcus faecalis have been characterized by oligonucleotide cataloguing of their 16S ribosomal RNA. All the organisms form a loose but coherent group that is phylogenetically equivalent to those of lactobacilli, bacilli, the Brochothrix and Listeria group, and related taxa that constitute one of several sublines within the ‘Clostridium’ branch of Gram-positive eubacteria. Within the Streptococcus–Enterococcus group, organisms fall into three moderately related clusters defined by Enterococcus, the lactic acid streptococci and streptococci of the pyogenic and oral groups, respectively.
-
-
- Short Communication
-
-
-
Maleness in Allomyces arbuscula
More LessColoured gametangia in the male and female hybrid Allomyces arbuscula × A. macrogynus and in various mutants of A. arbuscula were essentially of two sizes, large and small, measuring approximately 1847 ± 347 μm2 and 1032 ± 179 μm2 respectively. The large coloured gametangia were the same size as those of the female wild-type. The carotene content of the culture increased with masculinization and with increases in the proportion of large coloured gametangia. Relative gametangial size alone is not a reliable phenotypic marker of sexuality in Allomyces.
-
-
- Erratum
-
Volumes and issues
-
Volume 170 (2024)
-
Volume 169 (2023)
-
Volume 168 (2022)
-
Volume 167 (2021)
-
Volume 166 (2020)
-
Volume 165 (2019)
-
Volume 164 (2018)
-
Volume 163 (2017)
-
Volume 162 (2016)
-
Volume 161 (2015)
-
Volume 160 (2014)
-
Volume 159 (2013)
-
Volume 158 (2012)
-
Volume 157 (2011)
-
Volume 156 (2010)
-
Volume 155 (2009)
-
Volume 154 (2008)
-
Volume 153 (2007)
-
Volume 152 (2006)
-
Volume 151 (2005)
-
Volume 150 (2004)
-
Volume 149 (2003)
-
Volume 148 (2002)
-
Volume 147 (2001)
-
Volume 146 (2000)
-
Volume 145 (1999)
-
Volume 144 (1998)
-
Volume 143 (1997)
-
Volume 142 (1996)
-
Volume 141 (1995)
-
Volume 140 (1994)
-
Volume 139 (1993)
-
Volume 138 (1992)
-
Volume 137 (1991)
-
Volume 136 (1990)
-
Volume 135 (1989)
-
Volume 134 (1988)
-
Volume 133 (1987)
-
Volume 132 (1986)
-
Volume 131 (1985)
-
Volume 130 (1984)
-
Volume 129 (1983)
-
Volume 128 (1982)
-
Volume 127 (1981)
-
Volume 126 (1981)
-
Volume 125 (1981)
-
Volume 124 (1981)
-
Volume 123 (1981)
-
Volume 122 (1981)
-
Volume 121 (1980)
-
Volume 120 (1980)
-
Volume 119 (1980)
-
Volume 118 (1980)
-
Volume 117 (1980)
-
Volume 116 (1980)
-
Volume 115 (1979)
-
Volume 114 (1979)
-
Volume 113 (1979)
-
Volume 112 (1979)
-
Volume 111 (1979)
-
Volume 110 (1979)
-
Volume 109 (1978)
-
Volume 108 (1978)
-
Volume 107 (1978)
-
Volume 106 (1978)
-
Volume 105 (1978)
-
Volume 104 (1978)
-
Volume 103 (1977)
-
Volume 102 (1977)
-
Volume 101 (1977)
-
Volume 100 (1977)
-
Volume 99 (1977)
-
Volume 98 (1977)
-
Volume 97 (1976)
-
Volume 96 (1976)
-
Volume 95 (1976)
-
Volume 94 (1976)
-
Volume 93 (1976)
-
Volume 92 (1976)
-
Volume 91 (1975)
-
Volume 90 (1975)
-
Volume 89 (1975)
-
Volume 88 (1975)
-
Volume 87 (1975)
-
Volume 86 (1975)
-
Volume 85 (1974)
-
Volume 84 (1974)
-
Volume 83 (1974)
-
Volume 82 (1974)
-
Volume 81 (1974)
-
Volume 80 (1974)
-
Volume 79 (1973)
-
Volume 78 (1973)
-
Volume 77 (1973)
-
Volume 76 (1973)
-
Volume 75 (1973)
-
Volume 74 (1973)
-
Volume 73 (1972)
-
Volume 72 (1972)
-
Volume 71 (1972)
-
Volume 70 (1972)
-
Volume 69 (1971)
-
Volume 68 (1971)
-
Volume 67 (1971)
-
Volume 66 (1971)
-
Volume 65 (1971)
-
Volume 64 (1970)
-
Volume 63 (1970)
-
Volume 62 (1970)
-
Volume 61 (1970)
-
Volume 60 (1970)
-
Volume 59 (1969)
-
Volume 58 (1969)
-
Volume 57 (1969)
-
Volume 56 (1969)
-
Volume 55 (1969)
-
Volume 54 (1968)
-
Volume 53 (1968)
-
Volume 52 (1968)
-
Volume 51 (1968)
-
Volume 50 (1968)
-
Volume 49 (1967)
-
Volume 48 (1967)
-
Volume 47 (1967)
-
Volume 46 (1967)
-
Volume 45 (1966)
-
Volume 44 (1966)
-
Volume 43 (1966)
-
Volume 42 (1966)
-
Volume 41 (1965)
-
Volume 40 (1965)
-
Volume 39 (1965)
-
Volume 38 (1965)
-
Volume 37 (1964)
-
Volume 36 (1964)
-
Volume 35 (1964)
-
Volume 34 (1964)
-
Volume 33 (1963)
-
Volume 32 (1963)
-
Volume 31 (1963)
-
Volume 30 (1963)
-
Volume 29 (1962)
-
Volume 28 (1962)
-
Volume 27 (1962)
-
Volume 26 (1961)
-
Volume 25 (1961)
-
Volume 24 (1961)
-
Volume 23 (1960)
-
Volume 22 (1960)
-
Volume 21 (1959)
-
Volume 20 (1959)
-
Volume 19 (1958)
-
Volume 18 (1958)
-
Volume 17 (1957)
-
Volume 16 (1957)
-
Volume 15 (1956)
-
Volume 14 (1956)
-
Volume 13 (1955)
-
Volume 12 (1955)
-
Volume 11 (1954)
-
Volume 10 (1954)
-
Volume 9 (1953)
-
Volume 8 (1953)
-
Volume 7 (1952)
-
Volume 6 (1952)
-
Volume 5 (1951)
-
Volume 4 (1950)
-
Volume 3 (1949)
-
Volume 2 (1948)
-
Volume 1 (1947)
Most Read This Month
